Rebecca Calisi produced a must-see video (below) explaining the importance of bird research for understanding the brain. Please share it widely, because despite the obvious importance of bird research, there are still many who don’t get it, and misunderstandings and assumptions about bird researchers are rampant. As you will see, the video is geared toward those who are completely in the dark about bird research and those who conduct bird research.
We share so much in common with birds (ahem, evolution), and so our understanding of the brain, and especially hormone-brain interactions, has been shaped by bird biologists. Here are just a few examples…
The first evidence for hormone effects on brain and behavior were performed by Berthold in the 1800s; he studied roosters. He didn’t call the secretions hormones, but his work marks the beginning of endocrinology (the study of hormones, like those that control puberty, sexual desire, and the hormones in oral contraceptives and cancer treatments) and neuroendocrinology (the study of how hormones affect the brain and nerves in the body).
In the 1920’s Rowan discovered that day length (number of hours of light in a day) stimulates the reproductive hormones and behavior in dark-eyed juncos. This work initiated the field of seasonal biology and the effects of light and dark on mood, learning, depression, reproduction, and adaptation to seasons.
In the 1960’s Hinde and Lehrman opened an entire field of research based on their evidence that hormone-behavior relations are reciprocal, that is, hormones affect behavior, but changes in behavior affect hormone secretion. This research was on ring doves and canaries, and lead to the study of how our behavior and cues from our peers influences our own internal secretions (such as testosterone, serotonin, and dopamine).
Nottebohm and Konishi and their academic offspring made pivotal contributions to understanding how changes in brain cells (neurons) allow birds to learn songs of their own species, songs that they use to hold territories and compete for mating partners. This work with song birds is key to understanding how the brain learns and changes with experience including experiences with our sexual partners, experiences with stress and trauma (PTSD), and experiences with nurturing kindness from our caregivers.
Schlinger and Brenowitz discovered that rapid changes in the brain and behavior involved the enzyme, aromatase, in specific parts of the brain. This work with zebra finches and other species, is important for understanding the underlying cellular mechanisms involved when hormones have rapid effects on behavior, and when the behaviors of one individual affect the behaviors of another. It’s also important for understanding how brain cells survive trauma.
Again, these are just a few examples of why bird is the word.
Pictured, a Valentine, a birthday greeting, a bottle of wine, and a Jupiler
Jacques Balthazart is a true leader in behavioral neuroendocrinology, the study of how hormones affect the brain and behavior. His retirement demanded a fitting tribute, but there was a problem. When it comes to international conference organizers, nobody does it better than Jacques Balthazart. So, who would throw this party? No worries. The Belgian government’s policy requiring mandatory retirement at age 65 turned out to be the catalyst for a nonpareil scientific meeting (throughout this blog, all words underlined are links). It also turned out to be an outrageous birthday party and a creative plan to continue research on his own terms. Last week, the combination of foundational research, cutting-edge science, Belgian beer, and collegiality led to the conference’s new nom de plume: The International Conference Honoring Brilliant Balthazart (ICHBB).
Jacques is shown above and to the right just moments before the party, and below, just a few days into the party.
Photo by GianCarlo Panzica of Jacques Balthazart, 65, in a gift hat symbolizing his dual loyalty to Belgium and the U.S.
ICHBB Conference photos by GianCarlos Panzika
After all, when you are the premier conference organizer, entertainer, and hub of your scientific community, it makes sense that you should plan, host, and orchestrate your own birthday retirement party!
“Team Jacques” adapted from a funny photo by Julie Bakker
The first ICHBB was named the International Conference on Hormones Brain and Behavior and held in Bielfeld, Germany in 1982. It was conceived and developed from Jacques’ isolation as one of the few behavioral endocrinologists in Belgium. His uncontrollable desire for scientific interaction led him to invite about 40 premier behavioral endocrinologists from around the world to Bielfeld. To his surprise, they all showed up; it was an unqualified success; and everyone wanted to do it again and again. And again. In subsequent years, Jacques personally nursed the ICHBB in his home town of Liege (in 1984, 1989, and 2014) and affectionately nurtured the conference when it was hosted by others in France, Italy, and Spain.
I learned some things about Jacques’ life that I hope will be shared, remembered, and handed down to our academic offspring! First, necessity is the mother of invention in that some of Jacques’ biggest contributions to science come from his ability to embrace his authentic small-town lifestyle while uniting the world of behavioral neuroendocrinology. He was born, raised, educated, bred, and “retired” in Liege, Belgium, far less a tourist destination than a very pleasant place to grow up, and Jacques truly loves Liege. Many Americans have never even heard of Belgium, let alone, Liege, but one thing is very clear. Liege is “on the map” in the minds of behavioral endocrinologists. This just shows that there is no point in whining about where you work. I know behavioral endocrinologists at big U.S. medical schools, at Yale, or in big universities like UC-Berkeley who feel more isolated than Jacques.
Scenes from Liege
This idea was confirmed by plenary speaker, Kathy Olsen, former deputy director of the National Science Foundation, chief scientist for NASA, and associate director and deputy director of science in the executive offices of the President of the United States of America. According to Kathy, there is no one else to blame for putting Liege on the map other than Jacques Balthazart.
In terms of hormones and behavior, Jacques brought the world to Liege. I was surprised to learn that the man we know as the hub of our global science community is very much a local family man. Jacques’ father was a local architect, and his mother worked as a full-time homemaker and mother of two children. Jacques was educated from primary school on up through college in his home town of Liege. It was at the University of Liege that he fell in love with one of his biology professors, the beautiful and enviably fit, Claire (apparently she still runs 10K a day!). In addition to having a successful career in biology, Claire is a super friendly, social, community-oriented woman-about-town. Jacques, on the other hand, is not nearly as involved with his local friends and relatives. Though Claire chides Jacques for not remembering neighbors who have known Jacques his whole life, Jacques, ironically, is the social glue holding together a giant global network of scholars and friends, many of whom traveled far and wide to celebrate Jacques’ birthday. In one of Jacques’ presentations this week, he showed a world map dotted in every continent with markers showing where he has friends who will meet him at the airport.
Nicole, Lauren, Lauren’s son, Nancy Forger and Geert DeVries (left) and cannibals on the menu (below)
A Bad Bromance
Everyone in behavioral endocrinology knows that Jacques Balthazart has co-authored numerous landmark articles with Greg Ball, begging the question “How did this fertile collaboration begin?” During the various tributes at the conference, we learned that in 1983, Greg Ball was a graduate student at the Institute for Animal Behavior at Rutgers-Newark, where Jacques Balthazart was a distinguished visitor. Jacques, however, was not impressed with that Greg Ball character!
Jacques recalls Greg as a “lazy, long-haired hippie hanging around drinking coffee and pontificating in the break room all day long in a booming voice that could be heard all over the department.” As they say, first impressions are the best. Well, except the lazy part, because Greg’s and Jacques’ publications together number at least 400, and somewhere between 110-115 of those articles are co-authored by Ball and Balthazart or Balthazart and Ball.
The Ball and Balthazart Bromance was finally consummated (scientifically) a few years later in Germany. Greg Ball, then a postdoc with John Wingfield, was invited to speak at the conference. Greg was put up in a small dormatory-like room with a single bathroom shared by the adjacent room. Greg was brushing his teeth when his new next door neighbor, Jacques Balthazart, burst into the bathroom. “Well, well, well, we meet again!” Only this time, the Greg-Jacques Belgian beer bromance began in earnest (Wait? Who’s Ernest?) From the time of that meeting, Jacques Balthazart and Greg Ball became fast friends and insanely productive collaborators.
Incidentally, you can trace the academic family trees of these characters and that of your own mentor at Neurotree.org. Greg Ball’s tree is probably the most interesting, reaching straight back to Niko Tinbergen and Konrad Lorenz.
Peg McCarthy, Greg Ball’s beloved spousal unit, in her plenary lecture, explained that Jacques was a daunting obstacle blocking Greg’s affections for Peg in the initial stages of their courtship. At first, it was clear to Peg that Jacques would always be Greg’s “first wife.” It seemed she could never compete with Jacques. Luckily, Jacques came to adore Peg, and now warmly accepts Peg as a sisterwife. Suffice to say, that if all sisterwives were like Jacques and Peg, we would all be Mormons.
Greg Ball and Peg McCarthy (left) and a nicer pic of Peg with me and Colin Saldanha
Celebrating Belgium Moving Up In the World Cup 2014
To Sir Jacques With Love
The entire meeting was infused with enormous gratitude and affection from Jacques’ present and former students, postdocs, collaborators, family, and friends. Greg Ball gave a “What I learned from Jacques” speech that I wish all graduate students could hear. The highlights included
1) PUBLISH all of your data immediately. You never know when or how your results will be useful to other scientists, and none of it will matter if it is not published.
2) Time is precious, so, collect data, and write without fail, regardless of how late you stayed up the night before. No excuses. As Jacques would say, you have one 33 cl (Jupiler) at lunch and then bike back to work!
3) Be brave about methods. If it’s been done, you can do it.
4) Good ideas come from many sources, so, go to meetings and host your own.
5) Good colleagues can be good friends.
Jacques and Jeff Blaustein are not at all faking interest in my poster presentation. Photo taken by Vicki Luine.
In any case, there were so many excellent talks and posters at the meeting, demonstrating that Jacques will live on through his scholarship and mentoring as long as human civilization survives. Happily we learned that Jacques will continue working at the University of Liege, without the unpleasant duties of his old position, but instead intensifying focus on the research that he loves. This is one of the benefits of launching the careers of young, outstanding scientists and scholars. Shown below are ICHBB attendees who came to honor Jacques: Dave Grattan, Colin Saldanha, Kiran Soma, Jim Pfaus (with son, Josh), Thierry Charlier, Chuck Roselli and many others in the group photos.
Enjoy the next phase, Jacques! Your work is alive and well in all of our research programs. See you soon.
The above photo of Jacques and the preceding six were taken by GianCarlo Panzica.
There will be more pictures available through the website for the ICHBB. Meanwhile, you will find some links to seminal Balthazart discoveries here and here.
And finally, here’s a funky version of what I’m sure Jacques’ mentees are thinking…
This week the Schneider lab had the pleasure of hosting one of our science heroes, Paul Heideman from The College of William and Mary. He doesn’t wear a leather jacket or anything, and in fact, he looks like a typical white-guy professor. We’re still wondering exactly how he left us so inspired and energized, like we just discovered the thrill of science all over again.
Like most of us, Paul came up the ranks when everyone was advised to learn super cool state-of-the-art molecular techniques, to work on conditional knock-out/knock-in-optogenetic-whatever-the-hell mice, and to focus on cellular mechanism. Did he do this? No. Is he tenured? Yes. Is he funded? Yes, well funded. Is he happy? He seems pretty darn ecstatic.
Second, he studies the one thing that most biologists avoid like the plague: Individual differences. Most scientists seek to minimize them. They like groups that show little natural variance, and they omit outliers.
Hating individual differences makes sense in a way. When a drug works to reduce pain or fight infection, you want it to do so reliably in just about everyone. Will a new drug X decrease depression? We want a clean result. We want to see clear differences in depression between the drug-treated and placebo-treated groups. We want as little variation as possible within the groups. Face it. Too much variation within the treated group, and the drug is not going to market.
Vive la diffe’rence!
Paul solved the problem of individual differences by embracing them. He started his career in the field, trying to figure out why some equatorial animals are seasonal breeders and some aren’t. Fruit bats (a.k.a., flying foxes) that inhabit particular islands in the Philippines breed only in certain seasons, whereas those on other islands begin their breeding season months later. Presumably, this ensures that offspring are born at the time when they are most likely to survive. But what is the source of the difference?
To be a seasonal breeder, you need an internal clock, a reliable cue in the environment to set your clock (like the availability of food or the day length), and a sensory system to detect the environmental cue. Also, you need your clock and your senses to be connected to your reproductive system, and ultimately, to your gonads (ovaries or testes). What’s up with a nonseasonal breeder? Is the clock broken or are they deaf to the alarm? Is there a disconnect between the clock and the gonad?
Now What?
Paul was stuck. He just couldn’t figure this out in the field. In order to uncover the internal brain mechanisms, he needed to have animals in the lab. Commercially available lab animals, however, will not do. They gots no variance in the thing he wanted to study. Most lab animals are uniformly year-round breeders. He would have to create a lab population that had the same variation that exists in nature.
Being a rebel, he did what all of his colleagues were not doing. He trapped wild mice (Peromyscus leucopus) from the wild, brought them to his laboratory, and began crossbreeding. He kept the population large enough to avoid inbreeding (breeding close relatives tends to decrease heritable variation). Now he had a lab population that contained most of the variation that existed in the wild. How would he make use of this to answer his questions about individual differences in seasonality?
The next thing he did was brilliant. This base population served as the starting point for a selective breeding program. He started breeding lines of mice that differed in their seasonality. By breeding seasonal males to seasonal females, and unseasonal males to unseasonal females for many generations, he ended up with two groups of animals that differed from each other. They didn’t just differ by accidental inbreeding, or for other unknown reasons. They differed because they were selectively bred for those traits he wanted to study.
Paul has created and maintained a scientific gold mine. He can expose the two groups to winter conditions, measure their hormones and neuropeptide levels, and figure out what accounts for their differential response to the changing season. He can even sequence their genome and look for differences. He can get answers to questions like “How does evolution change the reproductive system?” “Can natural selection change hormone levels or does it change hormone receptors?” “In nonseasonal animals, is their internal clock broken or are they simply blind to the seasons?”
To find out the answers, you can check out Paul’s work here and here. Paul writes, “I have worked on multiple populations, but my current mice all come from one population. That’s important to me because I can say that all this variation is just from one little population — and that suggests that other mammals, including humans, might also contain large amounts of variation.”
I agree and I think it’s especially cool that selection has created wildly different strategies for survival and reproduction within the same species. In this case, you’ve got cautious mice that take the hint that winter is on the way by shutting down the reproductive system in order to conserve energy for survival in the cold. In the same population, you’ve also got flexible mice that will breed willy nilly as long as they can. If the winter is mild, the sexy mice beat the conservatives by producing litters and getting more genes into the next generation. If winter is harsh, the conservatives will win the gamble because they will be the only ones to survive to breed in spring. It takes all kinds. Vive la diffe’rence!
Paul gives a great talk. He gives all of his attention to the students, and he has tons of helpful advice about teaching behavioral endocrinology and science writing. Plus. . .
As Fat Tuesday approaches, my mind turns to cycles of indulgence and moderation. This is not news to women. They are quite familiar with regular, repeated bouts of uncontrollable appetite. Women are more prone to obesity and binge eating, and their binges are more likely to occur at a certain phase of the menstrual cycle. These differences are related, at least in part, to changes in hormones secreted from the ovaries. The ovaries secrete steroid hormones, such as estradiol and progesterone. Changes in estradiol and progesterone secretion alter the steroid environment in the brain and body, so that when hunger strikes, we might feel “just peckish” at one stage of the menstrual cycle or ravenously hungry at another phase.
How does this work? I received a four-year research grant from the National Science Foundation in 2013 to study ovarian hormone effects on appetite.
So, changes in hunger for food are correlated with changes ovarian steroid hormones, but correlations cannot tell you what causes what. My work started by looking for brain hormones secreted by cells with steroid receptors, brain hormones that increase the appetite for food and decrease the desire for sex. The problem is, we can’t really muck around in our own brains to study these hormones (neuropeptides). It’s difficult to study human food intake and sexual behavior because people lie about how much and what they eat. Don’t even get me started on measuring their sexual desire. No thanks. I like to study Syrian hamsters because I can precisely control what they eat, and they ovulate like clockwork every 4 days, unlike women who ovulate every 24-32 days. Plus, hamsters have a great way to demonstrate their hunger. After a period of dieting (say, we feed them only 75% of their normal daily food intake for a week), when we give them back their food, they increase their food hoarding. The hungrier they are the more food they carry in their cheek pouches from a distant source to their home cage. We can measure hamster sexual desire and hunger for food quite easily and accurately.
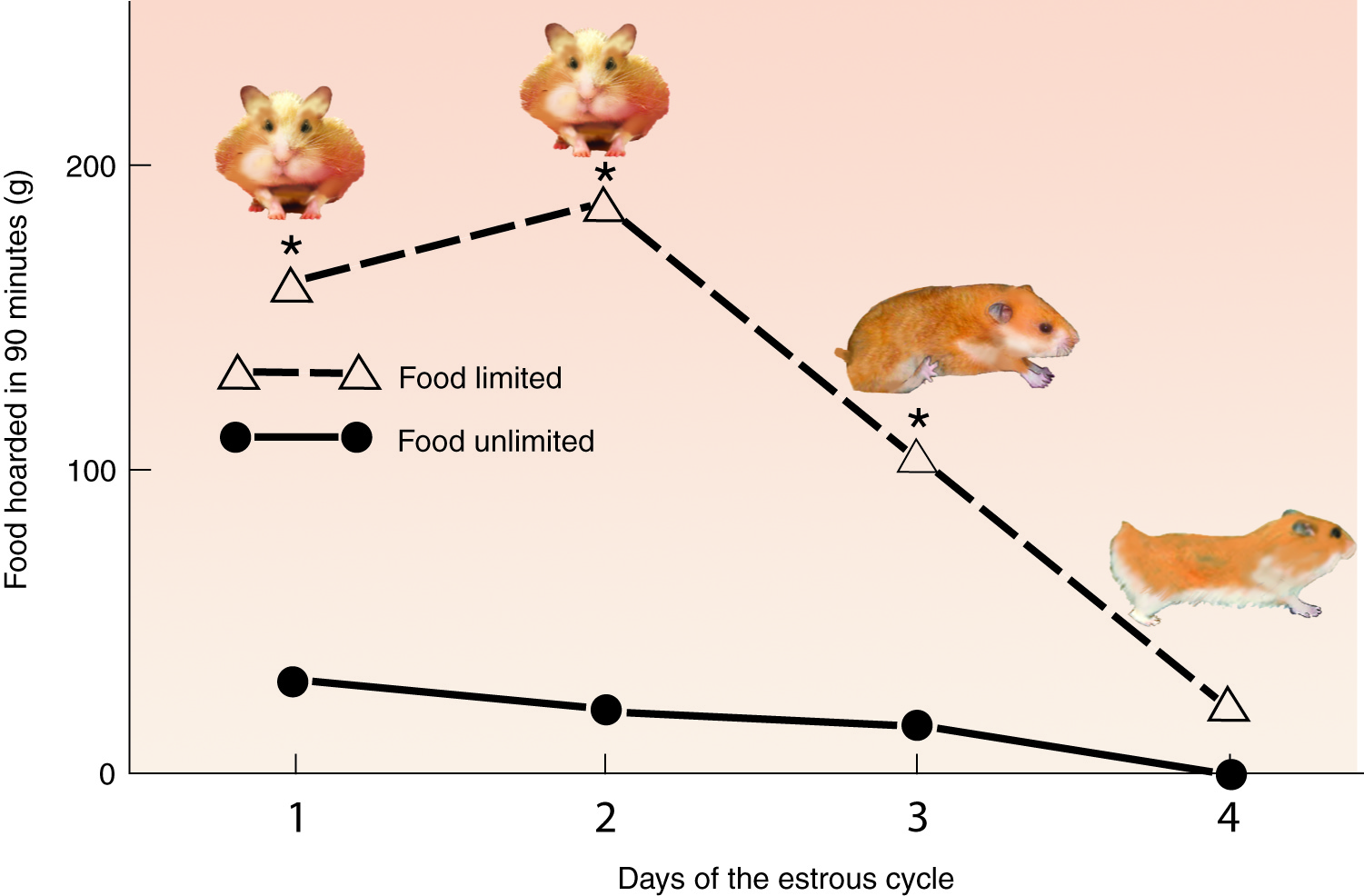
Hamster food hoarding (mean and s.e.m.) over the four days of the ovulatory cycle in food-restricted (open triangles, dotted lines) and food-unlimited (solid circles and lines) females housed with the choice between staying home, visiting a male, or hoarding food. The predominant sex behavior of the food-restricted female is shown in a cartoon above the hoarding data. On day 4 of the cycle, the periovulatory day, the females show mating behavior. On day 3, they show vaginal scent marking but do not mate. On days 1 and 2 they spend more time hoarding food than visiting the male. (Adapted from Klingerman et al., 2010 by Jay Alexander)
Candice Klingerman (a former grad student in my lab at Lehigh University and now a real professor at Bloomsburg University), found that hamsters on calorie-restricting diets show little interest in males and spend most of their time busily hoarding food, except on the day of ovulation. As ovulation approaches, however, they spend more and more time near the males. What you might find surprising is unrestricted females are obsessed with males throughout the ovulatory cycle! Whether they are ovulating or not, they ignore the food and spend more than 75% of their time leaving vaginal scent marks near the males. Like most rodents, they mate only on the day of ovulation, but the chubbier, calorie-unrestricted females prefer males over food every day of their four-day cycle. The differences between the calorie-restricted and unrestricted females are illustrated in the graph to the right, where you can see that calorie-restricted females do lots of food hoarding on most days of the cycle, with a conspicuous dip at the time of ovulation. The unrestricted females’ hoarding levels are low and flat throughout the cycle because they spend most of their time with the male (Klingerman et al., 2010).
You can see from the figure above that living in an “all-you-can-eat” buffet masks the effects of the ovarian cycle on the appetites for food and sex. Females on the “all-you-can-eat” diet consistently prefer to court males rather than stock their larder with food. Those females that are calorie-limited save all their sexual ardor for the small window of fertility on the day of ovulation. They spend the rest of the ovulatory cycle busily hoarding food. In the wild, this would ensure that there will be plenty of energy available for their offspring if their mating results in a pregnancy. This result makes me wonder whether our understanding of sex hormones has been clouded by studying animals housed in small cages with unlimited food. It makes me wonder how much our own species has diverged from our ancestors, now that we have adopted a sedentary lifestyle with food available in office vending machines, coffee break rooms, fast food restaurants, and well-stocked homes. No wonder we sit around watching Game of Thrones.
What are the brain differences between the hamsters on a limited-calorie diet and the hamsters at the “all-you-can-eat” buffet? I suspected that I would be able to find a brain hormone (neuropeptide) secreted by cells that have steroid receptors. I further suspected that the secretion of this neuropeptide is increased by food restriction. A review of the literature revealed many such chemicals. I have posted a handy table in a previous blog post here.
At the moment, we are interested in gonadotropin inhibiting hormone, GnIH. The figure below shows a hamster brain cell (neuron) that produces GnIH (a neuropeptide), which is stained red. Those GnIH cells that were activated by food restriction are shown in red with a green/yellow dot in the middle. These are cells labeled for GnIH and Fos, a marker for cellular activation. I got interested in GnIH when my colleague, Lance Kriegsfeld at the University of California at Berkeley, showed that GnIH inhibits reproduction in Syrian hamsters.
Brain cells stained for GnIH (red) and Fos (green). The red stain represents GnIH which occurs in the cytoplasm and thus colors a wide area of the cell body. The greed stain represents the proto-oncogene product Fos, a protein that is synthesized upon cellular activation. Fos resides within the cell nucleus. Cells that are red with a green/yellow stained nucleus are double-labeled with GnIH and Fos. These represent GnIH-containing cells that have been activated by food restriction. (Photograph and immunohistochemistry by Noah Benton)
Some of my other great colleagues in Australia (e.g., Iain Clarke) were showing that GnIH increases food intake in sheep, monkeys, and rats. GnIH sounded promising. Thus, I approached Lance about studying the effects of GnIH on the appetites for food and sex in Syrian hamsters. My student Candice Klingerman partnered with a grad student from the Kriegsfeld lab, Wilbur P. Williams. Together, Klingerman and Williams found that the level of calorie restriction was a good predictor of the level of GnIH cell activation (Klingerman, Williams, et al., 2011).
This suggested that GnIH might be part of the system that orchestrates the appetities for food and sex. This was confirmed by Noah Benton (Lehigh) and David Piekarski (UC-Berkeley). They administered GnIH to the brains of well fed females, and found that the GnIH-treated hamsters acted like they were starving. Their sexual appetites were lowered and their hunger for food was increased by GnIH treatment in the brain.
Another prediction you can make based on the hoarding data shown above is that GnIH will have different effects depending on the day of the ovualtory cycle (and the levels of estradiol and progesterone secreted from the ovary). Consistent with this idea, my student Noah Benton is finding that in food-restricted females, the activity of GnIH is elevated only during the nonfertile periods of the female cycle. In the figure above, GnIH cells are shown in red, and the activation of those cells is indicated by the central dot stained green for Fos, a protein that shows up in cells that have been activated. Noah double-labeled cells for both Fos and GnIH in food-restricted and food-unlimited females on every day of the ovulatory cycle. On nonfertile days of the cycle, there are significantly more GnIH cells activated in food-restricted compared to food-unlimited females. As you would predict from their sexy behavior, however, on the day of ovulation, GnIH is not elevated by food restriction. Go, Noah!
GnIH activity is usually elevated in food-restricted females, except at ovulation. We think the effects of GnIH are dampened by one of the hormones that is high around the time of ovulation. Noah Benton’s dissertation work will determine which ovarian steroid hormones and receptor are important for these effects. Will it be estradiol, progesterone, or testosterone? Place your bets.
Many obesity researchers think that appetite suppressing hormones are suppose to function to preserve our youthful figures and keep our body weights in fashionable and healthy limits. The work of my students shows that an important function of these hormones is to orchestrate the appetites for food and sex, perhaps to maximize reproductive success in environments where energy availability fluctuates. These effects are short-lived and change rapidly in the small time window of fertility (basically 1 day of the 4-day ovulatory cycle). It is probably unrealistic to expect any one of these neuropeptides to be a long-term or permanent cure obesity. Maybe we should think more broadly about how all this obesity has come about, and put some energy into understanding the link between energy balance (food intake, body fat storage, and energy expenditure) and reproduction.
“Sex and food and. . .” What pops into your mind to finish this sentence? For me it’s “rock and roll” (I substitute food for drugs because I don’t need drugs). The latter is also true for Seinfeld fans. Don’t Google “food and sex” though, you might get distracted.
“Sex-and-food” is the key to life. This grammatical impossibility stems from the fact that in the brain, the desires for sex and food seem to be one and the same, sometimes two sides of the same coin. In my upcoming book “Sex and food and. .” (Oxford University Press), I begin by noting that in every-day language food is sexy and sex is foodie;
“In both word and deed, we express our entangled appetites for food and sex, almost as if we confuse the two. In everyday language, food is sexy and sex is foodie. Chocolate is orgasmic and our lovers are delicious. In The Bible we are told to be fruitful and multiply. In Shakespeare’s Sonnet 118 he uses food metaphors for budding romance, for the “sweetness” of true love, and for the “bitter sauces” of infidelity.”
Just a smidgen of the evidence for the food-sex connection is presented in the table below. These 40-or-so chemical messengers have documented effects on ingestive behavior. They either increase or decrease the amount of food eaten per unit time. The same chemical messengers have clear, repeatable effects on reproductive processes, often including sexual desire and performance. In fact, compounds pushed as anti-obesity drugs by some researchers are being pushed as libido-enhancer by others. Our table reveals information useful for a wide range of scientists, not just those narcissists who feel that all research must have medical application. For more on this topic check out our new preprint now available online (it will be published open access).
agouti-related protein (AgRP), HS04, SHU9119 (MCR antagonists)
increases food intake in fish (Schjolden et al., 2009), birds (Strader et al., 2003), and mammals (Rossi et al., 1998; Stark, 1998), and food hoarding in hamsters (Day and Bartness, 2004)
inhibits gonadotropin secretion in fish (Zhang et al., 2012), inhibits LH in the presence of estradiol in female rats (Schioth et al., 2001; Watanobe et al., 1999), stimulates LH in male mammals (Stanley et al., 1999), ablation of AgRP gene restores fertility in ob/ob mice (Wu et al., 2012)
alarin
Increases food intake in male rats (Van Der Kolk et al., 2010)
stimulates LH secretion in castrated male rats (Van Der Kolk et al., 2010)
β-endorphin
increases food intake in fish (de Pedro et al., 1995b) (reviewed in (Lin et al., 2000)), birds (Deviche and Schepers, 1984; Maney and Wingfield, 1998; Yanagita et al., 2008), and rats (Grandison and Guidotti, 1977; McKay et al., 1981)
mediates stress-induced suppression of LH in fish (Ganesh and Chabbi, 2013), birds (Sakurai et al., 1986), inhibits LH secretion and sexual performance (Hughes et al., 1987, 1990; Sirinathsinghji et al., 1983), but might also increase sexual motivation in rats (Mitchell and Stewart, 1990; Torii et al., 1999),
galanin
increases food intake in fish (De Pedro et al., 1995a; Lin et al., 2000; Nelson and Sheridan, 2006; Volkoff et al., 2005) and rats (Kyrkouli et al., 1990)
stimulates LH secretion in birds (Hall and Cheung, 1991), steroid-primed rats (Sahu et al., 1987)
galanin-like peptide (GALP)
increases food intake in rats (Matsumoto et al., 2002), also decreases food intake in mice (Krasnow et al., 2003)
stimulates LH secretion in male mice and rats and in estradiol-treated female rats (Krasnow et al., 2003; Matsumoto et al., 2001; Uenoyama et al., 2008)
gamma aminobutyric acid (GABA)
might not mediate hyperphagic effects of orexin in fish (Facciolo et al., 2011); increases food intake in rats (Basso and Kelley, 1999)
increases gonadotropin release in fish (Kah et al., 1992); GABA-BR decreases excitability of mouse GnRH-I neurons (Zhang et al., 2009); GABA-AR excitatory for mouse GnRH-I neurons (DeFazio et al., 2002; Moenter and DeFazio, 2005)
melanin-concentrating hormone (MCH)
increases food intake in rats (Presse et al., 1996), but decreases food intake in fish (Shimakura et al., 2008)
inhibits LH secretion in rats (Tsukamura et al., 2000a)
neuropeptide Y (NPY)
increases food intake in fish (de Pedro et al., 2000; Lopez-Patino et al., 1999), frogs (Crespi et al., 2004), snakes (Morris and Crews, 1990), birds (Strader and Buntin, 2001), rats (Stanley and Leibowitz, 1984) and food hoarding in hamsters (Dailey and Bartness, 2009)
increases gonadotropin release in fish (Peng et al., 1993), inhibits steroid biosynthesis in frogs (Beaujean et al., 2002), inhibits sex behavior in snakes (Morris and Crews, 1990), inhibits LH in the absence of estradiol, stimulates LH in the presence of estradiol in rats (Crowley et al., 1985; Sahu et al., 1987) (Sahu et al., 1987), inhibits sex behavior in rats (Ammar et al., 2000)
orexin/hypocretin
increases food intake in fish (Lin et al., 2000; Volkoff et al., 1999; Volkoff et al., 2005), and rats (Sakurai et al., 1998), but not in birds (da Silva et al., 2008)
inhibits spawning in fish (Hoskins et al., 2008), inhibits LH in rats with little or no estradiol (Furuta et al., 2002), stimulates LH in rats with high levels of estradiol (Pu et al., 1998)
gonadotropin inhibiting hormone (GnIH)
increases food intake in birds (Tachibana et al., 2005), mice, sheep, and monkeys (Clarke et al., 2012; Johnson et al., 2007; Tachibana et al., 2005)
inhibits GnRH and LH secretion and sex behavior in fish (Moussavi et al., 2012), birds (Bentley et al., 2006; Satake et al., 2001) and blocks the LH surge in sheep and inhibits LH secretion in rats and female hamsters (Bentley et al., 2006; Johnson et al., 2007; Kriegsfeld et al., 2006; Smith et al., 2008)
Peripheral “Orexigenic” Molecules
Ingestive Effects
Reproductive Effects
corticosteroids
chronically elevated levels increase food intake in fish (Bernier et al., 2004), amphibians (Crespi et al., 2004), birds (Astheimer et al., 1992), and rats (Hamelink et al., 1994; McLaughlin et al., 1987; Stevenson and Franklin, 1970)
inhibits a wide array of reproductive parameters in fish including parental behavior (Carragher et al., 1989; O’Connor et al., 2009) reviewed by (Milla et al., 2009), inhibits steroid synthesis and spermatogenesis in amphibians (Moore and Zoeller, 1985; Moore and Jessop, 2003), inhibits sex behavior in snakes (Lutterschmidt et al., 2004; Moore and Jessop, 2003), stimulates gonadotropin secretion at low doses in birds (Etches and Cunningham, 1976), inhibits HPG function at chronically high doses in birds (Etches et al., 1984), and mammals (Vreeburg et al., 1988)
ghrelin (gut)
increases food intake in fish (goldfish and tilapia), but decreases food intake in rainbow trout (Jonsson, 2013; Jonsson et al., 2010), decreases food intake in birds (Kaiya et al., 2009), increases food intake in rats and mice (Tschop et al., 2000; Wren et al., 2000) and food hoarding in Siberian hamsters (Keen-Rhinehart and Bartness, 2005)
stimulates LH release from fish (Grey et al., 2010), inhibits GnRH, LH secretion and sex behavior in rats and mice (Fernandez-Fernandez et al., 2004; Furuta et al., 2001; Shah and Nyby, 2010)
insulin (pancreas)
chronically elevated levels increase body weight, adiposity, and food intake in birds (Nir and Levy, 1973), rats (Booth and Brookover, 1968; Friedman, 1977; Friedman et al., 1982; Houpt, 1974)
systemic treatment inhibits LH secretion at doses that increase food intake in hamsters not allowed to overeat (Wade et al., 1991), inhibits LH secretion in sheep treated peripherally with saline but not with glucose (Clarke et al., 1990)
motilin (gut)
increases food intake in fasted rats (Garthwaite, 1985)
inhibits LH secretion in rats (Tsukamura et al., 2000b)
progesterone (gonads, adrenals)
reverses the weight reducing effects of estradiol on body weight and food intake in rodents (Hervey and Hervey, 1966, 1969; Zucker et al., 1972)
synergizes with estradiol to stimulate female sexual performance in rats (Dempsey et al., 1936), enhances estradiol feedback on LH in female rats (Chappell and Levine, 2000), mimics testosterone in male rats (Witt et al., 1995)
testosterone (gonads, adrenals)
increases food intake and growth in rats (Siegel et al., 1981)
stimulates sexual motivation in females (de Jonge et al., 1986; Everitt and Herbert, 1970) and sexual performance in male rats (Davidson, 1966; Davidson and Bloch, 1969)
Central “Anorectic” Molecules
Ingestive Effects
Reproductive Effects
α-melanocyte stimulating hormone (α-MSH), melanotan- II (MT-II), PT-141
decreases food intake in fish (Kang et al., 2011; Schjolden et al., 2009), amphibians (Carpenter and Carr, 1996), birds (Kawakami et al., 2000; Tachibana et al., 2007), and rats (Vergoni et al., 1986), and food hoarding in Siberian hamsters (Keen-Rhinehart and Bartness, 2007a; Shimizu et al., 1989)
enhances electric organ discharge in electric fish (Markham et al., 2009), Stimulates LH secretion and sex behavior in rats (Alde and Celis, 1980; Thody et al., 1981)
Cocaine and amphetamine-regulated transcript (CART)
decreases food intake in fish (Volkoff et al., 2005), birds (Tachibana et al., 2003), rats (Kristensen et al., 1998)
stimulates GnRH secretion in rats (Lebrethon et al., 2000; Parent et al., 2000)
Cholecystokinin (CCK)
decreases food intake in fish (Himick and Peter, 1994; Volkoff et al., 2005), birds (Tachibana et al., 2012), rats (Gibbs et al., 1973) and food hoarding in Siberian hamsters (Bailey and Dela-Fera, 1995; Figlewicz et al., 1989; Teubner and Bartness, 2010)
stimulates GnRH and LH secretion in rats (Ichimaru et al., 2003; Kimura et al., 1983)CCK in the medial preoptic areas is required for estradiol-induced lordosis in rats (Dornan et al., 1989; Holland et al., 1997)
Corticotropin releasing hormone (CRH)
decreases food intake in fish (De Pedro et al., 1993; Matsuda et al., 2008), amphibians (Crespi et al., 2004), birds (Denbow et al., 1999; Furuse et al., 1997), rats (Heinrichs and Richard, 1999; Levine et al., 1983; Morley and Levine, 1982; Negri et al., 1985) and food hoarding in rats (Cabanac and Richard, 1995) reviewed by (Carr, 2002)
inhibits spawning in fish (Mousa and Mousa, 2006), inhibits LH secretion and lordosis in rats (Olster and Ferin, 1987) and sex behavior in Syrian hamsters (Jones et al., 2002)
Dopamine (DA)
decreases food intake in fish (Leal et al., 2013), rats (Heffner et al., 1977), increases food hoarding in rats (Borker and Mascarenhas, 1991; Kelley and Stinus, 1985), and reward (Wise, 2004)
inhibits gonadotropin secretion in fish (Omeljaniuk et al., 1989), stimulates sexual arousal, motivation and reward in birds (Cornil et al., 2005), rats and hamsters (Agmo and Picker, 1990; Meisel and Mullins, 2006)
Glucagon-like peptide (GLP-I)
decreases food intake in fish (Silverstein et al., 2001), birds (Tachibana et al., 2006), rats (Turton et al., 1996)
stimulates LH secretion (Beak et al., 1998)
Gonadotropin releasing hormone (GnRH I or II)
decreases food intake in fish (Hoskins et al., 2008; Nishiguchi et al., 2012), and female musk shrews (Kauffman and Rissman, 2004b)
stimulates LH secretion in fish (Moussavi et al., 2012), birds (Chowdhury and Yoshimura, 2004), stimulates LH secretion and sex behavior in amphibians and reptiles (Alderete et al., 1980; Licht et al., 1984), rats and sheep and sex behavior in shrews and mice (Kauffman and Rissman, 2004a; Kauffman et al., 2005) (Temple et al., 2003) (Moss and McCann, 1975) (Clarke and Cummins, 1982)
Insulin-like Growth Factor -1 (IGF-I in CNS)
ICV treatment decreases food intake in diabetic, but not normal rats (Lu et al., 2001), required for post-fast hyperphagia in rats (Todd et al., 2007)
restores LH surge amplitude in middle-aged rats (Todd et al., 2010), required for the LH surge, estrous behavior, estrous cycles in rats (Etgen and Acosta-Martinez, 2003; Quesada and Etgen, 2002; Todd et al., 2007), and for sex behavior in rats (Etgen and Acosta-Martinez, 2003)
Kisspeptin
decreases food intake in mice (Stengel et al., 2011)
stimulates GnRH and LH secretion in fish (Moussavi et al., 2012; Tena-Sempere et al., 2012), stimulates testicular expression of ER-a in frogs (Chianese et al., 2013), rats (Gottsch et al., 2004; Irwig et al., 2004)
Norepinephrine
decreases food intake in birds (Denbow, 1983) and stimulates food intake in rats (Ritter and Epstein, 1975)
inhibits LH secretion in rats (Iwata et al., 2011), stimulates sex behavior in birds (Cornil et al., 2005) and rats (Nock and Feder, 1979)
Oxytocin
decreases food intake in birds (Jonaidi et al., 2003), rats (Olson et al., 1991)
stimulates GnRH and LH secretion sex behavior in rats (Rettori et al., 1997; Whitman and Albers, 1995)
Secretin
decreases food intake in rats (Cheng et al., 2011a)
stimulates LH secretion in rats (Babu and Vijayan, 1983)
Serotonin (5HT)
decreases food intake in birds (Denbow et al., 1982), rats (Blundell, 1977)
stimulates LH in the presence of estradiol in rats (Coen and MacKinnon, 1979) Inhibits LH secretion in the absence of estradiol in rats (Coen et al., 1980) (Koh et al., 1984)
Thyrotropin releasing hormone
decreases food intake in rats (Vijayan and McCann, 1977) and Siberian hamsters (Steward et al., 2003)
stimulates LH secretion in pituitary in vitro not in vivo in rats (Fujihara and Shiino, 1983), and indirectly by effects on thyroid hormones in rats (Barrett et al., 2007)
Urocortin
decreases food intake in fish, amphibians, birds, and rats (Spina et al., 1996)
stimulates LH secretion in ewes (Holmberg et al., 2001), inhibits LH secretion in rats (Li et al., 2005; Nemoto et al., 2010), directly inhibits Leydig cell function in rats (Rivier, 2008)
Peripheral “Anorectic” Molecules
Ingestive Effects
Reproductive Effects
Adiponectin (adipocytes)
decreases food intake in rats (Bassi et al., 2012), increases food intake in mice (Kubota et al., 2007), decreases body weight and increases energy expenditure, insulin sensitivity, and ffa oxidation without effect on food intake in rats (Fruebis et al., 2001; Qi et al., 2004)
implicated in embryo implantation and fetal development in pigs and women (Palin et al., 2012), inhibits ovarian steroidogenesis in cows (Lagaly et al., 2008), inhibits GnRH and LH in rats and in GnRH cell cultures (Cheng et al., 2011b; Lu et al., 2008)
Adrenocorticotropic hormone (ACTH)
decreases food intake in rats (Vergoni et al., 1986)
stimulated LH secretion in female rats inhibits LH secretion in male rats (indirect via adrenals) (Mann et al., 1985; Putnam et al., 1991)
Bombesin (gut)
decreases food intake in fish (Volkoff et al., 2005), birds (Savory and Hodgkiss, 1984; Tachibana et al., 2010), and rats (Gibbs et al., 1979)
stimulates LH secretion in rats (Babu and Vijayan, 1983)
Cholecystokinin (gut)
decreases food intake in fish (Volkoff et al., 2005), birds (Savory and Hodgkiss, 1984), and hoarding in Siberian hamsters (Gibbs et al., 1973; Qian et al., 1999; Teubner and Bartness, 2010)
simulates LH secretion in rats (Perera et al., 1993); Inhibits lordosis duration in rats (Mendelson and Gorzalka, 1984), but see central effects in Table 1.1
Estradiol (gonads, adrenals, adipocytes, brain)
decreases body weight and food intake in fish (Leal et al., 2009), lizards (Shanbhag and Prasad, 1992), obese but not lean hens (Jaccoby et al., 1995; Jaccoby et al., 1996), rats (Nunez et al., 1980; Roepke et al., 2010; Roy and Wade, 1977; Zucker, 1969) and food hoarding in Syrian hamsters (Klingerman et al., 2010)
stimulates sexual receptivity and vitellogenesis and has negative feedback on LH in fish, frogs, lizards and birds (Chakraborty and Burmeister, 2009; Cheng, 1973; Crews, 1975; Gavaud, 1986; Gibbins and Robinson, 1982a, b; Licht et al., 1985; Liley, 1972; Mason and Adkins, 1976; McCreery and Licht, 1984; Redshaw et al., 1969; Shanbhag and Prasad, 1992; Yu et al., 1981), and stimulates LH surges in female rats (Chazal et al., 1974) and female sex behavior in rats (Dempsey et al., 1936; Powers, 1970), increases courtship and sexual behaviors in hamsters (Ciaccio and Lisk, 1973; Ciaccio et al., 1979; Takahashi et al., 1985)
Insulin (ICV treatment)
decreases food intake in rats and baboons (Chavez et al., 1995; Woods et al., 1979)
stimulates LH pulses in rats, pigs, and diabetic sheep and non diabetic ovariectomized sheep (Bucholtz et al., 2000; Cox et al., 1987; Daniel et al., 2000; Kovacs et al., 2003; Miller et al., 1995), inhibits LH in ad libitum-fed ovariectomized lambs (Hileman et al., 1993)
Insulin-like growth factor
increases body weight gain at superphysiological concentrations (Gruaz et al., 1997)
does not accelerate reproductive development in female rats (Gruaz et al., 1997)
Leptin (adipocytes, liver)
decreases body weight, adiposity, and food intake in fish (Crespi and Denver, 2006; Murashita et al., 2008), and mice (Campfield et al., 1995; Halaas et al., 1995; Pelleymounter et al., 1995) and food hoarding in Syrian hamsters (Buckley and Schneider, 2003) while increasing energy expenditure
mammalian leptin increases gonadotropin secretion in fish (Peyon et al., 2003; Peyon et al., 2001), delays the summertime regression of the testes in lizards (Putti et al., 2009), delays fasting-induced cessation of egg laying, follicular regression, and follicle wall apoptosis in chickens (Paczoska-Eliasiewicz et al., 2003), reverses the effects of metabolic challenges on gonadotropin secretion in mice (Ahima et al., 1996; Barash et al., 1996), estrous cycles, and steroid-induced sex behavior (the latter only in ad libitum fed female hamsters) (Schneider et al., 2007; Schneider et al., 1997; Wade et al., 1997)
Resistin
transient decreases in food intake in rats (Tovar et al., 2005)
promotes ovarian steroid secretion in rats (Maillard et al., 2011)
English: House mouse, 4 days old. (Photo credit: Wikipedia)
Mind blown. I just discovered that data I collected one afternoon during my junior year of college is important and useful, now. I was sitting at the banquet of the annual SBN conference, when former president of the society, Art Arnold, said “Hey, that paper you published 30 some years ago has been so useful in getting researchers to start paying attention to sex differences.” At first I was confused. Was he talking about that old review article I co-authored with my postdoctoral mentor (which keeps getting cited even though we’ve both written much better reviews)? Or maybe he meant the overrated article we published in Science in 1989? No, it turned out he was excited about a paper that emerged from a side project loosely related to my FSU undergraduate honors thesis in which I measured the length of mouse taints.
It all started when some grad students and a postdoc decided they wanted to give the googly-eyed, work-study student (me) a “discovery experience.” This, by the way, is the essence of mentoring, and I am forever grateful to those guys. They gave me 50 or so one-day-old mouse pups and said “Figure out which ones are male and which are female; collect some data; and show us how to document the sex difference.” I sat alone in the room, like the miller’s daughter in Rumplestiltskin, staring hopelessly at the seemingly identical embryonic bodies (see pic). I focused on the place where you might expect to see a tiny penis, but where I hoped to see a penis, there was only a small bump and a small hole behind the bump. Each and every mouse had the same bump-and-hole arrangement. Finally, after more staring, I noticed that the bump-to-hole distance was small in some mice and large in others. The hoity toity science name for this is the “anogenital distance.” To the likes of me, the term for this distance is the taint, because it tain’t the genital and it tain’t the other thing. So, I divided the pups into two piles, a short-taint pile and long-taint pile. Now how was the miller’s daughter going to spin mouse taint into scientific gold? My Rumplestiltskin was a professor who studied the visual system, the late Howard Baker. I told him I was trying to measure a very tiny distance that I could barely see with my naked eye. He gave me a reticle, a glass eyepiece for a microscope with a ruler engraved on the lens, which allows measurements accurate to 0.01 mm. I measured all the little mouse taints, and found the mean for each group. I did a t-test, and the difference was highly significant. I guessed that the those mice with the smaller taints were the females. Correct! Chuck and John, the grad students and postdoc in the lab already knew the answer, but since I discovered it without any help, they encouraged me to publish a paper. Hence the 1978 publication in Behavioral Research Methods and Instrumentation, “Determining the sex of neonatal mice, Mus musculus.“
Howard Baker never demanded my first-born son, few colleagues since have been so generous regarding authorship, and I rarely think of this one-page publication in a minor journal. Half the time I don’t even list it on my c.v. I just assumed in 1978 that everyone knew the importance of knowing the sex of your experimental animals on the day of birth. Tragically, the majority of scientists stubbornly refuse to look at both sexes in their experiments. The fact is, males and females differ in response to pain, drugs, hormones, and their propensity for many different diseases. Sex hormones have profound effects during early development, both pre- and neonatally, and these hormones masculinize or feminize the individual, determining their adult response to drugs, cancer, infections, pain, diet and exercise. We now have a large body of data on common diseases and biological processes; most of it on males by a ratio of 5:1. In 1993, the importance of sex differences and of early hormonal effects prompted the NIH to mandate the enrollment of women in human clinical trials. This mandate should also apply to the animal research on which the human research is based. Excuses for the male-only bias include “females are too variable due to their estrous cycles,” and even more ridiculous, “I don’t know how to tell the difference between the sexes.” It taint rocket science; even a work-study student at a state school could figure it out.

 Rebecca Calisi produced a must-see video (below) explaining the importance of bird research for understanding the brain. Please share it widely, because despite the obvious importance of bird research, there are still many who don’t get it, and misunderstandings and assumptions about bird researchers are rampant. As you will see, the video is geared toward those who are completely in the dark about bird research and those who conduct bird research.
Rebecca Calisi produced a must-see video (below) explaining the importance of bird research for understanding the brain. Please share it widely, because despite the obvious importance of bird research, there are still many who don’t get it, and misunderstandings and assumptions about bird researchers are rampant. As you will see, the video is geared toward those who are completely in the dark about bird research and those who conduct bird research.


 Balthazart. So, who would throw this party? No worries. The Belgian government’s policy requiring mandatory retirement at age 65 turned out to be the catalyst for
Balthazart. So, who would throw this party? No worries. The Belgian government’s policy requiring mandatory retirement at age 65 turned out to be the catalyst for 
































{kind=link}